
Introduction
The idea that Ecology is a young science was brought home to me when I gave a lecture at the Open University in 2019 as part of the 50th year anniversary of the University’s foundation in 1969. My own 50 years of ecological research has sought to clarify and refine a range of ecological concepts so that more appropriate measurements can be made and from there mathematics applied, predictions made and theories tested. Like many ecologists, I and others, seek in this way to mature the young science of Ecology.
My Open University lecture sought to provide a context for the 50 years of Ecology’s early days before 1969 and examine the progress made in the 50 years during the OU’s existence to 2019. I had gone to work there in 1972 as a recent graduate from Durham University and what struck me in preparing my lecture was that the 50 year span of the Open University was very long when compared to the active scientific history of Ecology itself.


Of Ecology’s founding fathers the one that most exercised me was one Raymond Lindeman. Lindeman’s paper in 1942 further developed Tansley’s concept of ecosystem to establish the mainstream perceptions of Ecosystem of today. Lindeman less successfully suggested that ecosystem evolution was driven by maximising the trophic level efficiences of food webs. That paper was just a 30 year old when I had started at the OU in 1972. Now half a century later it’s coming up to 80 years old and still in my view, leaving key ecological concepts unresolved.
Ecosystem
This is the most important concept in Ecology but in other ways the most distracting and hardest idea to pin down. It is the ‘system’ part of Ecosystem that causes the problem. This can be illustrated by the question, What is a HUMAN SYSTEM? Then your choice is wide indeed. Is it a factory, a house, an amusement park, a hospital, a city? If we instead ask what object is a human, then a human being is pretty clear choice and we know roughly how many there are on the planet. Humans are countable.
So what is the candidate for the Eco’system’ object?
It is an energy catchment, or photonshed, analogous to a watershed, where solar energy enters the food chain via sunlight and passes through plants and animals until some of that energy reaches the top predator. Top predator territories create separate photosheds, hence establish separate ecological objects. These I argue are the largest entities that can be made using ecological processes. There are larger objects on the earth, such as lakes or rivers, but these are part of the global hydrological cycle and in the case of rivers, run across top predator territories or in the case of lakes, have aquatic top predators making one or many ecological objects within the lake, or sea, or ocean.
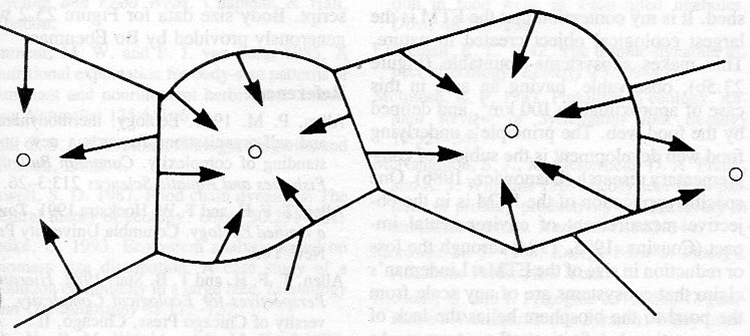
Some ecologists have taken these ideas seriously, I am glad to say. The Encyclopedia of Ecology and Environmental Management edited by Peter Calow (1998), details my approach in its Ecosystem entry.

More interestly, especially from the long term history of Ecology, is Leibold and Wootton’s republication (2001) of Elton’s 1927 founding classic, Animal Ecology, where they take the opportunity to review the 70 year progress of Elton’s ideas. Here they note that my process based top predator definition of community is in sympathy with Elton’s approach rather than the current preference for area based communities/ecosystems of lakes, forests etc. They make the analogy of my process based definition of ecosystem with its potential for advancement to Mayr’s (1942) process-based definition of the species concept.
It is conventional to view the ecosystem via energy and material flows rather than as population dynamics. But as Martinez (1995) neatly put it, cells use energy and materials and pass these across their boundaries, single organisms and therefore populations do likewise. There is in this sense no special ‘energy level or currency’ separate from the organisms that constitutes an ecosystem; there are only organisms.
Ecosystem Size
Humans as top predators and human impact
Humans definitely have been and in some cases, still are, group top predators when hunting prey with tools and as such would have defined the food web boundaries to create the ecosytem object. As with lion determined top predator areas, these are the largest things that ecological processes can make until humans began trading between local clans created larger integrated structures for the exchange of goods. If lions could have traded a range of goods they may have created village markets, market towns and cities on Earth where exchange of goods and ideas takes place. This hierarchy of human settlement is well known to geographers and arises out of the economics of trade. From this perspective the human population, once it leaves direct predation on wild food, it no longer defines the ecosystem object, in that sense trading humans are not part of the ecosystem but leave it and are part of the economic system creating towns and cities at a vastly larger scale than the ecological object. The great American urbanist Jane Jacobs (1992) was the first person I knew who explicitly bought into this view!
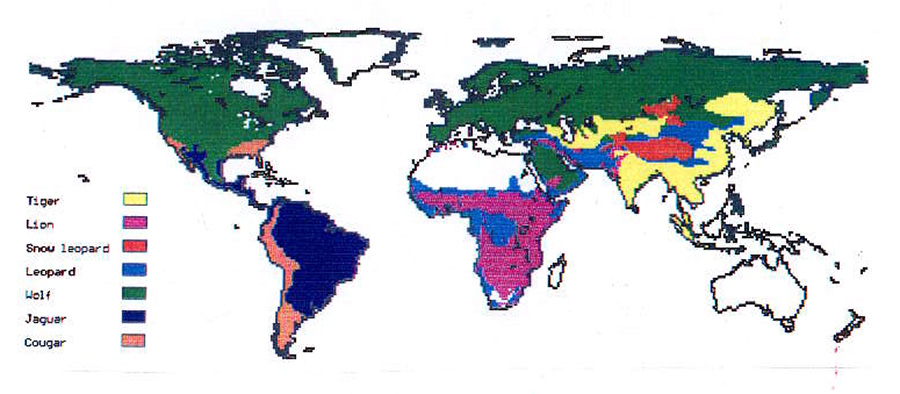
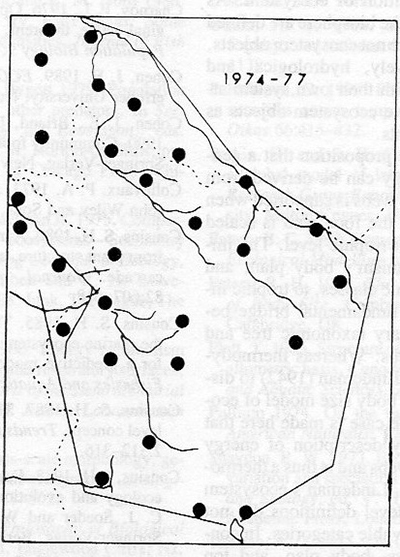
Using this approach human impact on the ecosystem can be measured by counting how many full sized top predator territories still exist and compare that to estimates of pre-human dominance. For Lions in Africa we estimated a continental population of 1,100,000 animals and 78,000 living in 1980, i.e. representing 7% of ecosystem objects at that date. These are estimates of individuals rather than group territories, prides.
Our idea at the time was to do a global count of all full sized top predators and compare to the primitive distribution to assess a total human impact. This led to work on Wolves globally as one other species.
Hierarchy Theory
This is the part of ecological science that tries to make sense of what are the ‘parts’ in ecology and indeed also the ‘parts’ relevant to evolution since we are told biology only makes sense in the context of evolution. So what are the parts and how do they relate to each other.
One of the really nice things about the ecosystem being defined by top predators is that these structures can evolve because the predators can evolve and they have continuity through time by virtue of having offspring which is an essential feature (Hull 1974).
Keeping things brief, it is important to distinguish between objects that define themselves (self-organising entities) such as organisms which are there whether we measure them or not and ideas that we impose on the world such as trophic levels which are abstract and are dependent on us humans to create in our minds.
One exciting idea coming from hierarchy theory is that for parts to have properties they have to be independent (at least to a significant degree) from their own parts and the things that they are part of. That independence is achieved by storage. Thus plant based terrestrial ecosystems are locally independent of climate by water retention in the soil – known to climate modellers as ‘the bucket’ – and the organisms in the food web are independent of each other by virtue of fat stores. Thus expenditure of energy by an organism does not have to be replaced, that second, by eating something and as a result the organisms have independence and behaviour.
Food Webs
There are many authors who point to food webs, food chains and all things trophic as being at the core of Ecology. So for example, my defining the Ecosystem object from food web properties clearly fits with this. As McNaughton et al (1989) put it neatly, ‘Ecosystems are structurally organised as food webs’.
Food and Feeding therefore require careful measurement and careful modelling if we want a rational ecological science.
There is still a fundamental divide between Elton’s body size analysis of food webs and Lindeman’s condensation of feeding relationships into trophic levels; plant, herbivrore, carnivore1, carnivore2. The appeal of this attempted rationalisation laid the foundation for the 10 year International Biological Programme 1964-1974. Indeed it was my good fortune to attend the wrap-up conference for IBP at The Hague in September 1974. The decade long programme of applying trophic level techniques to all of the Earth’s major ecotypes did not provide the expected viable outputs or synthesis. But did boost ecology as a ‘big science’ which is perhaps it major beneficial outcome. For me though I wanted to understand why trophic level analysis (TLA) had not worked and what could replace it.
There are many flaws in TLA but one is that the green plant is treated as a Unity to scale trophic space. Seeds, leaves, wood are very different food items yet bundled as trophic level 1 thereby scaling everything that eats these different items as TL2. Elton did not do this and he noted different size based food chains coming from different parts of the plant/tree.
The other more fundamental problem is that TLA is a historical model of energy flow; each trophic level has the same number of historical energy transfers from the plant. Yet we know that feeding interactions are determined by the present state of the energy availability not its history. Imagine that the coins in your pocket could be isotope tested to show that this penny had been spent 4 times and that 6 times since issue, the pound coins 2 and 4 times, then sort the money into piles of coins that had been spent the same number of times. Try decoding the economy based on peoples purchasing behaviour based on coins that had been spent 4 times, 6 times and so on. Its pretty clear that in economics too the value of the coin trumps it’s history, although of course the coins will have had a history – its just not relevant. In Ecology we need to know the value, i.e. quantity and energy availability, of the food item to judge its properties in the ecosystem. The trophic level system does not do this and hence fails as a model and did fail in IBP’s most exhaustive test. Knowing the food history via isotope analysis does not help; nor will it help the predator make it’s decision on what to kill/eat. But the lure of the TLA is very strong in a true Paradigmic manner. My New Scientist article was an attempt to scupper the TLA and for a few years it seemed to have an effect. But now the paradigm is back ironically while body sized base trophic analysis has bounced back strongly too.
So what to do?
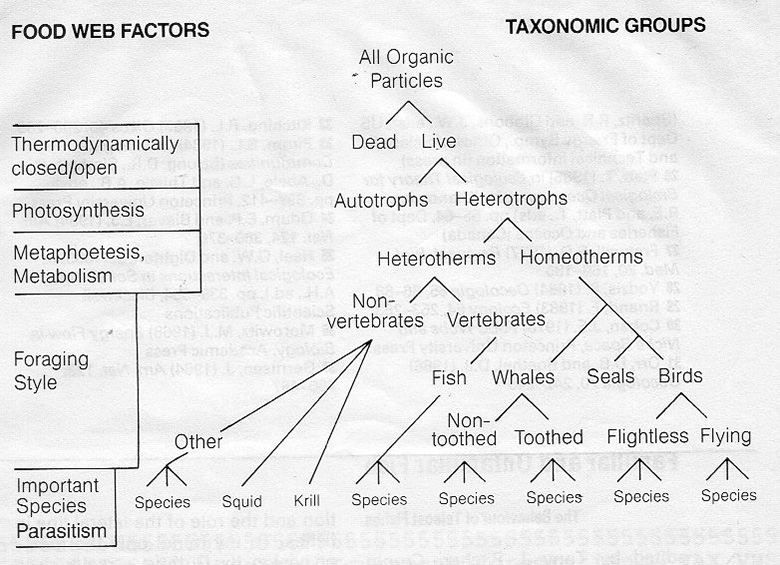
The obvious thing is to create different models of feeding interaction. These should recognise the variation in energy availability in the green plant and have feeding interactions between animals that are based on present state nutritional value, not historical status. Detritus could be similarly modelled as it moves to oblivion. There would be no levels, it would be a continuum, but a distinction made between the dynamics of the plant, reproducing animal populations and non-reproductive, indeed decaying carcasses and detritus. The notion of different groupings in the model with different dynamics (3 mentioned in this high level case) and sized based interactions between predator and prey. In the image below a predator of a given size is feeding on a range of smaller prey. This work from 1981 and 1985 has been confirmed by Williams and Martinez (2000).
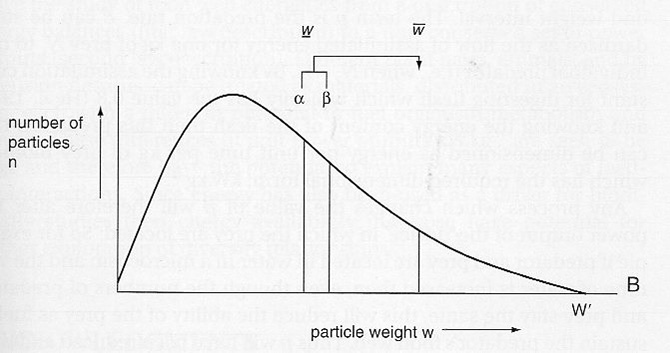
The predator itself will grow in size until adult changing its prey ‘window’ as it grows. Metaphoetesis (Lindeman’s supervisor Hutchinson’s term) is the change of diet accompanying the growth of the predator – imagine Lion reproduction involved starting life as thousands of bee sized individuals few of which get to full size – but fish do this. In reality different taxonomic groups have different foraging abilities arising from different body plans, birds, bats, molluscs and so on. These can be separated out according to model objectives taking into account different functional food web factors.
Biodiversity
Seemingly this is the easiest of our ecological concepts, not least because organisms are countable and different species are often easily separated in observations. This makes it a good proxy for observing the state of habitat and how that might change over time. However when we want to use biodiversity as part of a body of theory or for conservation purposes then it becomes much more opaque. Species are by definition different to each other. We are familiar with the idea that you cannot add apples and pears yet the maths of diversity indices certainly does that and does not differentiate between an eagle and a sparrow for example 1+1=2 in a species count. The Shannon-Wiener index is for example very sensitive to the number of species in a sample which in turn is a function of the sample size. There are now several ways of allowing for differences between species both functionally and for the purposes of conservation.
General Theory of Ecology
To mature ecological science we need to have clear insights into our phenomena in general and food webs in particular, especially if we invite members of other disciplines such as maths and physics to come and play in our sandpit.
I have previously tried to synthesise my own journey in a paper given at the Colarado Pingree Park Food Webs conference in 1993. I reproduce the final paragraph of that paper here.
In Conclusion
“In conclusion, the proposition that a general theory of ecology can be derived from the operation of food webs is supported when the trophic space is scaled by body size and not trophic level.
The importance of an organism’s body plan, and therefore of taxonomic classes, to trophic interactions creates a fundamental bridge between the evolutionary taxonomic tree and function in food webs. Whereas thermodynamics was used by Lindeman (1942) to displace Elton’s (1927) body size model of ecosystems, the opposite case is made here that body size is the key description of energy availability in food webs and is thus a thermodynamic description.
Lindeman’s ecosystem and higher trophic level definitions did not create directly observable categories. In contrast, organism mass, body plan, and top predator territory form an observable basis from which to create a coherent ecology structured on the food web. Defining the ecosystem from the food web gives ecology intrinsic scale and differentiates ecology from other sciences.“
Further Information
Contact
For further information please contact Steve Cousins
Steve Cousins has a BSc in Zoology from Durham University and a PhD from Open University where he worked from 1972 to 1989 and was a Research Fellow in the energy research group. From 1989-2006 he was Associate Professor at the Honda International Ecotechnology Research Centre, Cranfield University.